
Stochastic corrector
Eörs Szathmáry's "Stochastic corrector" model is seen by
many as making the error catastrophe problem faced by early
organisms less pressing.
Here I discuss the relevance of this model to the origin of
life.
Motivation for the model
Szathmáry's model deals with a population of distinct
self-replicating entities in dividing cells.
It deals with the problem of how information could be
transmitted to offspring - in the form of the ratio between
the constituent replicators - despite the fact that these
replicators are not physically joined together - thus:
- Stochastic forces present when the cell divides
distributing them unevently in offspring;
- Differences in the rate of reproduction of each sort of
replicator within the cell;
Explanation of the model
The model is best explained with a diagram:
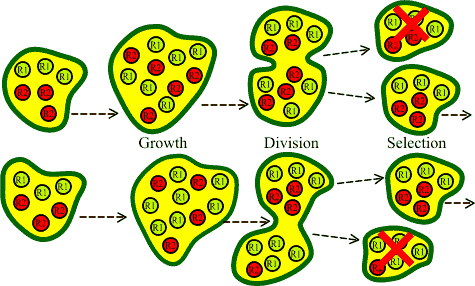
Organisms are composed of a small number of different types
of self-replicating agents: R1 and R2.
The organisms work best if there is some specified ratio
between the replicators: in the diagram the preferred ratio
is 1:1.
The cells grow and eventually split into two - with the
replicators being distributed randomly between the offspring.
Then selection acts - destroying any cells that deviate too
far from the optimal ratio of replicators within each
cell.
The result is that the information content represented by
the replicators - and their proportions in the cell - is
preserved between generations.
The model can still work - even if some of the
replicators reproduce faster than other ones - and thus
there is between-replicator selection within each cell.
Good points
The model is useful - since it shows how a small collection
of small molecules - none of which may be capable of
independent replication could co-exist in a community - and
help catalyse each others replication, and be inherited
without being physically connected together or
having their division orchestrated by some sort of controller.
Not so good points
However the model doesn't scale up very well - the more
replicators are involved the greater the chance
that stochastic variations will result in one type of
replicator being omitted from any children - and the
stronger selection is needed to maintain verbatim
transmission of information between the generations.
Rewriting the model
I think the model makes a lot more sense if it is rephrased
a bit:
Rather than consider the diagram as representing different
sorts of replicator in a cell, consider it as representing
different sorts of replicator in an ecosystem:
For example, if you have a "pool" filled with replicators,
then the species in it may colonise another pool downstream.
If a key species gets wiped out - or fails to get
transmitted to the new environment - then the new ecosystem
will not flourish.
The idea that the proportion of replicators in each
"cell" is somehow significant is abandoned in this model.
That information is no longer strongly inherited.
However, the collective genomes of all the important species
still get transmitted to the next generation - i.e.
information about the existence of the different
species is preserved.
This phrasing retains a key feature of the model - namely
the possibilty of inheriting a lot more information
than is present in any individual replicator.
It also still leaves open the possibility of a community of
mutually-dependent organisms surviving in an environment where
none of them could exist alone.
Overcoming differential reproductive rates
What about the possibilty that one sort of replicator will
wipe out the other ones?
Szathmáry's original model invoked a kind of group selection
to explain how the system could overcome this problem -
suggesting the selection between the ecosystems would
act to penalise ecosystems where one replicator does
too well.
Selection between ecosystems is likely to have been an
important factor.
Some other points may also be worth mentioning, though:
- Independent niches
- If the different replicators do not compete
significantly for resources then they may be able to co-
exist peacefully - without one wiping the other out.
Niches might be partially independent in an ecosystem -
the replicators in the system:
- ...may have different nutrient needs;
- ...may exist in different areas of the ecosystem.
- Frequency-dependent selection
- It's in all the replicators interests to prevent the
population of any one replicator from dropping too low -
since the loss of an essential player is terminal for
everyone involved.
This fact tends to favour cycles involving
frequency-dependent selection - where the
least-numerous critical replicator is
preferentially synthesized.
- Mutual dependency
- It's possible that one replicator may depend
on the products of another one to successfully reproduce.
Such dependencies may produce frequency-dependent
selection that favours the scarce replicators.
The membrane-free corrector
This reformulation no longer depends in any way on the
notion of a membrane or cell - instead, the role of
container could equally well have been played by the
environment - which could have been as simple as a rock
pool.
Membranous material is unlikely to be involved in the
earliest living systems - since the organic material that
composes most membranes tends to form sticky messes - that
are incompatible with the process of crystallisation that
is likely to be responsible for the replication of the
genomes of the first organisms.
Notes
The idea presented here owes an obvious debt to the model
presented in my earlier Increasing
Complexity essay - i.e. it is basically the same
idea wrapped up in different terminology.
References
- Eörs Szathmáry and J. Maynard Smith - The Major Transitions in Evolution. Oxford, 1995.
- Eörs Szathmáry and J. Maynard Smith - The Origins of Life, Oxford University Press, 1999;
- Eörs Szathmáry and László Demeter - Group selection of early replicators and the origin of life. Journal of Theoretical Biology 128, 463-486, 1987;
tim@tt1.org | http://originoflife.net/
|